Falling Larynxes and Noisy Neighbors
Evolving Creative Minds II
Just-so stories, unfortunately, comprise the bulk of what passes for explanatory discourse in evolutionary psychology, including its applications to human creativity. Often interesting in their own right, they are best understood as elements of a pre-scientific method of coming to terms with complex historical data.
Why It's Not Just So
Such stories begin by adopting the adaptationist program as Gould and Lewontin, 1979 would have us understand it, namely swallowing the postulate that traits we observe in organisms are there because they help spread copies of the genes associated with them; in other words, traits are adaptive. Natural selection, the prime mover of evolutionary change, winnowed them from among the competing options. To understand specific structures, functions or behaviors we accept that they are adaptations and try to guess what it is about them that could help genes spread.
But must they adaptations? Of course not. Our look at micro- and macroevolution and the physics of ontogeny has been it plain that any number of "forces" are continuously at work in shaping biological diversity and organismic design. Natural selection is one among these. Its primacy needs to be carefully evaluated for each case with data illuminating the comparative roles of all elements of the evolutionary mechanism. To do otherwise risks falling into fallacies of unwarranted extrapolation. That, plus two other traits of just-so stories to be considered next, is why adaptationist discourse can spark such debate, even among evolutionists.
A common adjunct of just-so storytelling is the extreme genetic reductionism based on the erroneous formula "gene ---> trait" (compare "change in gene" ---> "change in trait, maybe", discussed earlier). Individuals do not duplicate themselves during the process of reproduction. They replicate their genes and then scatter them into the future. If a gene blueprints a trait, then it is easy to portray individuals as passive, gene-programmed vessels for polynucleotide replicators. All traits of an organism become nothing more than enabling devices for the expansive replication of hereditary material. As we saw above, differential replication of gene variants is a universal property of biological evolution, but not because one gene equals one trait or because all the genes taken together make up a blueprint in which the organisms resides homunculus-like.
Like New Testament parables, just-so stories instruct and entertain. We will meet some of them below. As scientific objects they are best regarded, I would like to suggest, as the work (not the best work) of a young science on its way to discovering how to make testable predictions about biological history. In the hands of contemporary masters of the genre (scholars such as Dawkins (1976) and Wilson (1978), for example) they rivet attention on key adaptive issues while stimulating debate. They are, however, no substitute refutable hypotheses, i.e. conjectures which not only rationalize what is known but stick their necks out to predict, and be beneficially smashed apart in consequence.
What We Know
We are a young species, and our species "family," its genus, is also a new kid on the block - barely 5-7 million years old, a moment in macroevolutionary time. A few hours in a tenure hearing or a faculty promotions committee will remind even the staunchest sapiens-phile that we remain fully primate in the major parameters of our social behaviors as well as in our anatomy and physiology. Seen against the macroevolutionary backdrop afforded by the all the social species currently known (Wilson, 1975 is still the incomparable reference here; see also Trivers, 1985 and Tudge, 1996), we are not pushing the primate "envelope" all that hard.
Genetically we are very close to our nearest living relatives, the chimpanzees, given what superficially seems our physical and cognitive differences from them. In fact so close are humans and chimpanzees in details of anatomy, physiology, chromosome structure, biochemistry and gene sequence that they resemble many pairs of animal species such as fruit flies and birds known to have split from each other only during the past million years! In his recent overviews of human evolutionary, to which the interested reader can turn for much more background than will be needed here (see also Tattersall, 1993, and references therein), Jared Diamond (1992, 1995) notes that recent studies put the fraction of our genome in which we actually differ from the common and the pygmy chimpanzees at about 1.6%, a number that falls to under 0.2% when allowance is made for the portion of this minute difference that is likely to be so-called "junk" DNA (free-rider polynucleotide text with no known function).
Such small numbers are remarkable, but they need be surprising only in view of an extreme genetic reductionism that seeks to draw a straight line from genes to the "finished" organism. You will recall from the discussion of nonlinear dynamics in development that even minute inputs (changes to the system) can cause big results by nudging it across a threshold, triggering a big change - a burst in brain size or voice box redesign, for example. In the nonlinear world 0.2% or less can go a long way if conditions are right. No empirical evidence I know of currently supports a conjecture that such a nonlinear bifurcation separates us from our primate and hominid cousins. The idea is interesting though.
The pattern of physical evolution expressed by the exquisite array of prehuman fossils and artifacts suggests that creativity's fingerprint on our history will need careful interpretation. In a word, things seem to have been very slow for creativity for a very long time in the ancestral paleoworld. Hominids were upright and bipedal by around 4 million years ago. Some 2 million years later Homo habilis was abroad in East Africa and hominid brain size was increasing (650 cc in round figures; compare about 400 in the chimp, 450 in Australopithecus africanus, 1200-1400 in modern humans; Tobias, 1971, 1979). A burst in cranial capacity lasted from the follow-on Homo erectus at 1.5-1.7 million years (900-1000 cc, roughly) through to around 100 thousand years ago, when it leveled off with the appearance of the first anatomically modern Homo sapiens in southern Africa.
So, 5 million years ago we were, so to speak, still sitting in the trees; 3 million years later we were standing upright and using simple but effective stone tools; almost 2 million years after that we were still using stone tools, albeit of greater diversity and impressive refinement. Along the way fire got harnessed and some of us eventually began burying our dead, accompanied perhaps by rituals of pre-interment bodily adornment. But judged purely on the basis of the physical evidence, a quarter-million generations of hominid creativity had changed our invented world about as much as the handicrafts made by a modern child change the content of their closets in the dozen or so years between kindergarten and college.
Then, about 40-50 thousand years ago, in western Europe, 50 thousand years after the appearance of people with brains as big as ours and skeletons hooked together just like the ones we have, material creativity took off: specialized and compound tools, fabricated dwellings, long-distance trade, the masterpieces of cave art sleeping through the millennia at Lascaux and its kindred forgotten sanctuaries. The rest is history.

Our radio telescopes, tuned ever more astutely to the whispers of the night sky, might one day bring us contact with some ancient, spacefaring species who walked the rift valleys in hominid times, so that in their exo-anthropology libraries we will at last see the behaviors and hear the voices of Homo habilis and Homo erectus. Until then we must return to the fossils and the mute stone implements, interpolating as best we can, tuned to their inherent biases rather than the buzz of intragalactic static.
These biases are especially important to paleopsychologists reconstructing creativity's slow evolutionary dawn because our data are limited to what history gives back to us. Like soft bodied life forms that live suspended in warm seas and perish without leaving more than an occasional, tantalizing wiggle in the ooze of the ocean floor, many conceivable outcomes of the creative process will leave the physical record untouched. At least, the traces will be hard to detect with a scholarship focused, as ours currently is, on recovery and debate about inventions, rather than the far more difficult task of inferring the existence of a created work from its impact on what does gets made and discarded - second- and higher-order effects, if you wish. If you paint with colored sands your masterpiece is gone with the next wind. The singer of new variants of Homeric songs within the oral tradition may live to see those variants dispersed and blended as they become the mental property of other oral masters.
Dolphins Do It Too
We can turn to the creative activity of other species as well as to our own culture, with its frenzied pace of just-in-time obsolescence and the Next Best Thing, to see this ephemeral face of creativity. Without the patient field work of Japanese primatologists (Kawai, 1965), the invention of potato washing by Imo, the macaque genius, would leave traces only in terms of the what a later fossil record would show of population densities, living sites, migration routes. Could we deduce potato washing from that, with our present methods?
Wet mammals, as well, are rusing to take second place to their dryer primate cousins. Bottlenose dolphins Tursiops truncatus, a species for which there is some evidence of self awareness (Marten & Psarakos, 1995) along with obviously big brains and a standing reputation for playfulness, have more recently come up with a novel behavioral innovation at Sea Life Park in Hawaii (Marten, Shariff, Psarakos & White, 1996). Though lacking hominid prehensile capacities to fashion material objects, the Sea Life dolphins have discovered how to use their blowhole apparatus to fashion delicate but stable air bubbles in the shapes of large rings and helices (a Quicktime movie currently is available at www.sciam.com). Air and swirling water are their raw materials.

The skills needed to make even the simplest forms of air-filled rings require practice, and Marten et al. report that the behavior is socially transmitted in the sense that novice dolphins acquire ring-making skill in the presence of experienced ring makers. Laka, a female dolphin who is an especially innovative ring master, has discovered many other novelties. She has been observed releasing air bubbles from her mouth and flipping them into the ring once they have passed along her body, and adding air to a ring by exhaling more into it. These behaviors are monitored closely by other dolphins, who will sometimes coordinate their own ring making with them. Similar behaviors have also been reported from other oceanaria and among cetaceans in the wild.
Creativity, like evolution, is opportunistic in is release. On your next walk along an ocean beach the rolling waves will not speak the achievements of Laka or those who came before her. Theirs is an invented world of ephemera. Dolphins do not write, and bubbles do not fossilize. Simil the minds of Homo habilis, Homo erectus, and the earliest Homo sapiens have vanished along with all but the most obvious and enduring traces of their creative activity, including their genetic residuum in us. So the data available are biased (one is perhaps justified in saying almost hopelessly biased) toward the least ephemeral signs of their minds' best work. Methods are needed for sensing the traces that innovative ephemera leave on such fossil and paleocultural assemblages. Like invisible writing brought out by waving a time-eaten letter over a candle flame, the message of creative ephemera must, if possible, be added to the story of inventiveness recorded in the stone tools.
True primates, we are fascinated by novel things we fashion with the coordinated action of our eyes, hands and brain to spin our invented world of texts, gadgets and gizmos, things we make to please, amuse, seduce, and our worlds. It is important not to underestimate the role in our evolution of innovated social ephemeral like the earliest chants and narratives (Kreindler & Lumsden, 1994). Our childhood encounters with them build our minds. There can be no serious evolutionary theses about creativity until this paleoarcheaology of ephemera has been developed to its limits and integrated fully with the data on "endurables," and the field work on play and innovation in animals integrated with human data.
Suppose, however, that such future work produces a story of creativity's evolution in the genus Homo more or less consistent with what we now know, so that the small groups of habilis and erectus were as short on sharable ideas, tales, ceremonies and chants as they were on types of implements as they huddled against the Pleistocene night. We then return to face the troubling discord between a fossil record that shows a rush toward big brains, agile hands, and bipedal locomotion but a long, slow climb in the press of creativity out into the world - a crawl that ended with a burst of material innovation about 400 centuries ago.
Falling Larynxes and Noisy Neighbors
There are some good stories told by evolutionists to reconcile the phylogeny of artifact diversity with the notion of Darwinian adaptation. These synthetic tales are worth looking at because they show alternative patterns in which the evidence, briefly introduced above, can be marshaled in support of specific functional interpretations. Properly developed, these interpretations might someday reach a point at which they will give novel, refutable predictions. Let us consider a few examples.
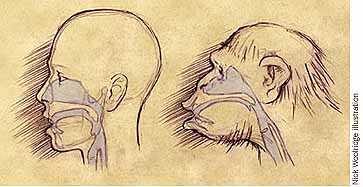
Jared Diamond (1992, 1995), whose recent considerations of human paleobiology I have referred to, has suggested that we can understand the mismatch between the run of brain size and inventive diversity in the following way: hold back about 0.01% of our genome and suppose that changes in it put spoken language into its modern form, about 40 millennia ago. Creativity, boosted by the recombinant powers of language and linguistic cognition in their modern forms, blasted off.
Using primate and anthropological examples, Diamond infers that before the hominid mind took hold of modern language we were basically somewhat smarter chimps, getting on utterances akin to human pidgins and creoles. Changes in that bit of our genome renovated our vocal tract and hooked it properly into our already enlarged brains, with its ready-and-waiting processing power. We could now communicate more effectively about novel ideas and, perhaps, equipped Fodor-like with a more powerful "language of thought," slice and dice ideas in our heads, the better to come up with fresh ideas in the first place. The 60 millennium stretch between early sapiens and the creative explosion tuned up our ability to utter vowels and consonants, and the way our brains did complex grammar, allowing us to leave erectus and neanderthalensis in the dust.
Perhaps. But what held back that critical 0.01% for so long? One small step, after all. And can we be confident that the complexity of modern human grammatical capacity was a necessary condition for Upper Paleolithic artifact diversity? Is language really a sine qua non for creativity in general? What about adaptive opportunities provided by more creative approaches to social interactions (Cheney and Seyfarth, 1990; Cosmides & Tooby, 1992) or courtship, mate competition, and matters of sexual selection (Trivers, 1985)? Language only, or something more? Was there really an inventive stasis prior to the Upper Paleolithic, so that a burst-like, punctuated-equilibrium metaphor is appropriate to the data?
Perhaps diversity was increasing in the usual way all along (e.g. Tobias, 1979, Fig. 14). Many evolutionary trends in innovation follow exponential curves, for example, or logistic curves whose initial behavior is essentially exponential (Hamblin, Jacobsen and Miller, 1973 is a nice compendium of examples from the time period in which the neoDarwinian synthesis was being created). The early part of an exponential curve can look very flat, and appear so if sampled at coarse resolution. Once the slope of the exponential (which is always increasing) and the magnitude of the curve (which is also always increasing) rise above our method's level of resolution we get a signal, like a blip suddenly appearing on a radar screen. Coarse sampling can make a gradual pattern look like a jump
John Pfeiffer (1982) ties the Upper Paleolithic's cognitive and creative upswing to broader patterns of change involving population growth and steadily increasing pressures on the people and the land. Put crudely, for perhaps the first time in human history necessity became the true parent of invention and creative outcomes found increasingly receptive audiences: more people, more hungry and harassed people with a need, more people to spread the news, more places to spread the news to. Creativity made innovations that allowed people to make more people faster, further increasing the pressure for innovation. (Perhaps the creative potential had always been there, dormant, awaiting necessity's cold bite.) The gene-culture connection locked in and the "modern" period of rapidly intensifying creativity in human history erupted. The load of cultural information pressing down on ever more complicated networks of band societies, where people could not yet read or write, opened new opportunities for people adept at drawing; the underground sanctuaries at places like Lascaux became socialization machines in whose bowels rhythmic ceremony, sensory deprivation and exposure to virtual worlds compellingly rendered on the cavern walls helped ingrain band norms and wisdom.
Perhaps. It helps to have an audience, though even a Leonard Bernstein or a Garth Brooks needs to press CDs if they are playing to a future specialized in tracking endurables rather than the murmurs of ephemera. Or, perhaps, creativity was not stalled at some uninspired pseudochimp level or triggered by noisy neighbors, but moved steadily from pattern to pattern as hominids moved down out of the trees and steadily toward skyscrapers.
Merlin Donald (1993) has suggested that our records of both Homo erectus and the early Homo sapiens bear the signs of major changes in cognition and culture linked to creativity. The breakthrough for erectus, supporting its (eventual) transcontinental journeys and harnessed fire, was vastly improvement voluntary motor control involving the whole body, not just the vocal tract, serving mimesis and allowing them to reenact, share, and plan - to create what Donald calls a "gestural prototheater of everyday life". The second breakthrough is similar to the changes suggested by Diamond, involving reorganization of the vocal tract and attendant neuroanatomy, higher-speed speech skills, and the breakthrough to one of the more potent channels of ephemera creation: lexical invention and the proliferation of vocabularies supporting planning and narrative-based enculturation (Kreindler and Lumsden, 1994).
Perhaps. But do we know that kinematic aping was any more important than creolic vocalization in coming to terms with the challenges faced by erectus? Human creativity is intentional and the activity of a self-aware mind. Where is consciousness and how is its evolution tied to the story of creativity? Perhaps the earlier adaptive need was for self-awareness first and foremost, the better to outwit game and rival suitors. Perhaps this transformation of consciousness was well along by the Upper Paleolithic and completed its journey as an unexpected spin-off of, say, vocal tract redesign.
Creativity is a flirtation with complexity, the risks of change bearing against imagined triumphs. Neural processes serving such a capacity must, at the very least, be able to pack and shuffle a lot of information in the tight space of a hominid cranium already occupied with other duties. Perhaps human creativity does hang suspended in history by a thread spun from less than 0.2% of our DNA, but maybe the critical change was not voice or language, but a delicate elaboration in the pattern of cortical wiring to, which burst the Paleolithic bandwidth for splicing ideas and testing the their merits. Maybe it was a need for better sleep to ward off the stresses of band life that favored neural rewiring for subtly altered REM, with a burst in dreaming and creativity (e.g. Feldman, 1988; Kavanaugh, 1996). Evolution can be like that, one thing triggering another in unexpected, even preadaptive cascades. Perhaps.
Wonderful speculations like those offered by Diamond, Pfeiffer and Donald can be multiplied indefinitely. They are part of the circum-scientific mythology by which we try to make sense of ourselves. Even the best ones, such as the examples visited here, ingenious in their formulation and valuable in the unity they bring to diverse data, are guesses cast in the adaptationist mode. Too soft to refute as they stand, they induce more than they deduce. But that, I believe, must hardly be considered the point. Evolution is a young science of nature's most complex creations; if we are to write an evolution of creativity that merits the label scientific, future work must understand how to test the validity of the adaptationist stance itself as well as deductive models placed within it, deconstructing both relentlessly when needed to unearth creativity's true phylogeny.
COG to the Rescue
The evolutionary science of creativity does not exist. One day it may, but the subtle nature of the mental phenomena at issue and the spotty nature of the historical makes the task formidable, perhaps impossible. In other words, irresistible. I would therefore like to suggest that the following need to be kept in mind if we are to move our understanfing of evolution and creativity closer together:
- It is clear that something happened in the Upper Paleolithic. Not so clear is whether this is the first poke of a long-preexisting signal above background, or an evolutionary breakthrough, or something else yet again. As data improve it may be possible to more rigorously test "smooth diversity" and "punctuated creativity" hypotheses against the evidence, and thus against each other.
- Quantitative modeling may be increasingly helpful in tying numerical counts of artifact categories to creativity. A good evolutionary hypothesis should be able to predict the mathematical form of the diversity curve in times like the Upper Paleolithic explosion. An unbroken line of smooth exponential growth, for example, may be consistent with a gradual progression joining the creativity of physically modern humans across to early Homo sapiens and Homo erectus. A punctuation model would imply at least a change in rates from one predictable range to another.
For example, in information-propagation models based on the concept of "percolation," information moves from site to site in a lattice of information processing and transmission units (Stauffer, 1985). Below a certain threshold of conductivity among the units (which might be neural circuits in the brain or foraging bands roaming a plain) signals cannot find complete paths through the system and input gives no output. But above the critical threshold information is processed through the system. The mathematical form of the system response in the vicinity of the threshold falls into a restricted number of categories called universality classes. To what universality class does the Upper Paleolithic transition belong? This datum limits the range of allowed evolutionary and paleoecological processes consistent with the transition.
- Does creativity compute? Reaching down into the depths of the quantum universe, Roger Penrose (1994) proposes that human creative thinking is noncomputable. This flagrantly iconoclastic assertion negates all conventional thinking in neuropsychology and cognitive science, which takes mind as grounded in the physics of neural cell interaction. While perhaps exceedingly complex, these interactions can be studied in numerical terms akin to any other physical process.
Although I am not aware of any cross- species comparisons, the human mind clearly is at least a Turing machine capable of intentional, universal computation up to the limits permitted by our finite memory capacities (see also Tudge, 1996, p.253). The universal Turing machine or UTM sits atop a hierarchy of abstract mathematical devices that define progressively more powerful kinds of information processing (introduction and review in Dufort and Lumsden, 1997). The point where the hominid mind attained capacities akin to the UTM ranks as a phylogenetic milestone. But Penrose would have us go much further than this, arguing from his study of mathematical invention that human creators can innovate in a way no computable process ever can. It is very important to realize that this is not a statement on bounds caused by finite resources like the numbers of transistors or the amount of memory. It is about the limits achievable by following algorithms, however large or complex. You will recall that algorithms or their equivalent are the basis of all mathematical approaches to the mind, including the ones used so far in computer models of creative work. If Penrose is on target (Dennett, 1995, among many other defenders of the traditional doctrine, states the reasons why most people think Penrose is way off track), all of these efforts are wrong because creativity is not computation, and somewhere in evolution, likely on the branch leading to us, appeared noncomputable minds with our creative phenotype.
- Computability may create. In total contrast to the Penrosian position is the hypothesis that creativity can be understood precisely because it is an algorithm (Dasgupta, 1994; Langley et al., 1987). It can be written out and put in a computer. Since at least the time of Campbell's seminal paper on variation and retention (Campbell, 1960), evolutionary metaphors have stimulated psychologists' reasoning about the human creativity (Gruber & Davis, 1988; Perkins, 1994, 1995; Simonton, 1993; Sternberg & Lubart, 1995). Digital incarnations like BACON (Langley et al., 1987) and AARON (McCorduck, 1991) take the next step. By following algorithms they do seemingly creative work in complex domains such as physics (BACON) and artistic drawing (AARON). Perhaps our vaunted creativity is a lot simpler than we'd like to suppose. Penrose gets stood on his head. But Douglas Hostadter (1995) has issued a cautionary
Current creativity machines are disappointingly domain-restricted and dependent on abundant up-front hints to make their alleged creations and discoveries. More exotic software architectures (Hofstadter, 1995; Minsky, 1986) may, however, achieve true creative work, giving evolutionary scientists a new logical calculus for framing hypotheses about paleocognition. Most promising in this regard is Rodney Brooks COG project at MIT, which is attempting to grow (I can think of no better term) a mind akin to a young child's inside a social robot.
The frenzied debates COG has sparked among cognitive scientists are documented elsewhere. The point here is that if the ethical dilemmas about playing with the development of potential (quasi-)persons can somehow be resolved, even modest success with COG will revolutionize the paleopsychology of human creativity, simply because COG's learning rules can be redesigned at will, and its learning environment shaped to match anything from Lascaux to a Homo habilis campsite.
- Great minds matter, but how much and for whom? Creativity research has richly plumbed the effusia of a few dead minds (Bloom, 1994; Boorstin, 1992; Gardner, 1993; Kearney, 1988; Perkins, 1981; Petroski, 1994; Simonton, 1988). Our myths laud the Homers, the Newtons, the Michelangelos. Geniuses and heroes all. But we know too little about the influence such individuals exert on social history (Csikszentmihalyi, 1988), let alone on evolutionary history (Findlay and Lumsden, 1988). In the play of contingency and catastrophe, what (or who) matters (Shermer, 1993)? Chaos theory has made the butterfly effect into a modern parable: nonlinear systems like societies and ecosystems can get into "chaotic states" in which they are exquisitely sensitive to small effects (Lansdown, 1991). The weather is a popular example: the butterfly flapping it wings in an Amazonian rainforest (or Laka blowing a ring bubble in Hawaii) sets off the hurricane that trashes the Gulf coast of Texas. Not always of course; the conditions must be poised just so. Would our creative great ones have dominated culture regardless, or did they happen to be flapping their wings at the right moment in history? Is this the butterfly effect in action? What was its role in prehistory and the evolution of creativity?
Ghosts in their Shell
Sitting with us on our branch of the evolutionary tree are numerous other species for which play is a key mechanism of learning and skill rehearsal in the young. We can, as well, chart the ebbs and flows in creativity for human lifetime. But evolutionary biologists have scarcely begun to assimilate these life history patterns and place them into the context provided by other species. This is a deficiency, because we saw above that evolutionary science is driving for a new unification with developmental biology; we cannot explain intact organisms through genetic reductionism.
Take the developmental concept of neoteny, for instance, the persistence of traits present in the young into adulthood (review in Raff, 1996). The globe-pated, snub-muzzled layout of the human skull resembles that of young apes more than it does the cranial topography of adult apes. In a classic instance of an ontogenetic just-so story, genetic changes in the ancestral primate development program delay circumorbital and related enlargements in the human skull and jaw, giving us our flat-faced approach to the world. A glib spin on human creativity would trace its origins to a neotenal delay that keeps play and the desire for novelty, exploration, and experimentation, in many species the prerogative of the young, open through adulthood. No doubt there is more to be said.

Evolutionary biologists are, in addition, beginning to follow the lead of psychologists (Gardner, 1992; Sternberg, 1985) in recognizing intelligences specialized for specific domains such as language, visual expression, or social interactions (Cheney & Seyfarth, 1990). "Creativity" is still too often treated as a monolithic penchant for variation and selection. Did creativity take off because hominids had an adaptive opportunity a one domain, like language? Was that diversity engine then appropriated by natural selection to serve creativity elsewhere, or has human creativity evolved more than once in specialized ways, to deal in the separate contexts of language, art, and society?
A reasonable question asks what can keep future thinking about puzzles such as these from being just a superabundant stock of adaptationist tales under the subject heading, "How We Got Inventive." I see no alternative to demanding of evolution what we demand of any science historical or otherwise, namely conjectures that can be tested in the hard light of new data. There is no reason at present to suppose that such tests will cast natural selection from its place as a key shaper of biodiversity and organic form. But better methods will give us a more precise understanding of just when and where adaptation has been important, and how it has worked in concert with the other processes of evolution to shape the creative mind and its self-spawned niche, culture.